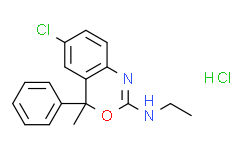
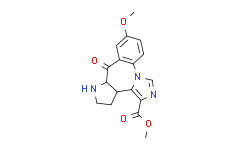
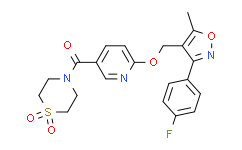
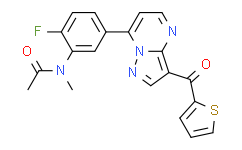
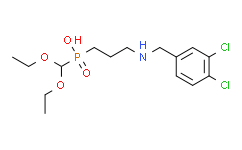
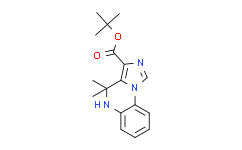
- 精细化学
- 生命科学
- 分析化学
-
Signaling Pathways(信号通路)
- Anti-infection(抗感染)
- Protein Tyrosine Kinase/RTK(蛋白酪氨酸激酶/RTK)
- Apoptosis(细胞凋亡)
- Autophagy(自噬)
-
Cell Cycle/DNA Damage(细胞周期/DNA损伤)
-
DNA/RNA合成
-
PARP
-
HSP(热休克蛋白)
-
Sirtuin
-
ROCK
-
p97
-
PLK(Polo-like Kinase)
-
Topoisomerase(拓扑异构酶)
-
PERK
-
Antifolate
-
Casein Kinase(酪蛋白激酶)
-
ATM/ATR
-
HDAC(组蛋白脱乙酰基酶)
-
Deubiquitinase(DUB)
-
Chk(Checkpoint Kinase)
-
Aurora Kinase
-
RAD51
-
TOPK
-
Wee1
-
Nucleoside Antimetabolite/Analog
-
SRPK
-
IRE1
-
PPAR
-
CDK
-
G-quadruplex
-
Eukaryotic Initiation Factor
-
Mps1
-
DNA-PK
-
APC
-
DNA Alkylator/Crosslinker
-
Haspin Kinase
-
PAK
-
DNA/RNA合成
- Cytoskeleton(细胞骨架)
- Epigenetics(表观遗传学)
-
GPCR/G Protein
-
Prostaglandin Receptor(前列腺素受体)
-
mAChR
-
Adrenergic Receptor(肾上腺素能受体)
-
Sigma Receptor
-
GHSR
-
Histamine Receptor(组胺受体)
-
Angiotensin Receptor(血管紧张素受体)
-
Adiponectin Receptor(脂联素受体)
-
Bombesin Receptor(蛙皮素受体)
-
Ras
-
Glucocorticoid Receptor(糖皮质激素受体)
-
Cannabinoid Receptor
-
Adenylate Cyclase(腺苷酸环化酶)
-
GPR40
-
Neurokinin Receptor
-
GPR139
-
Dopamine Receptor(多巴胺受体)
-
CCR
-
Leukotriene Receptor(白三烯受体)
-
CXCR
-
Melatonin Receptor(褪黑素受体)
-
Neuropeptide Y Receptor
-
LPL Receptor
-
mGluR
-
Adenosine Receptor(腺苷受体)
-
P2Y
-
PAR(Protease-Activated Receptor)
-
GPCR19
-
GPR119
-
GPR84
-
Urotensin Receptor
-
GPR120
-
Endothelin Receptor
-
Opioid Receptor
-
CaSR
-
CGRP Receptor
-
Cholecystokinin Receptor
-
CRFR
-
EBI2/GPR183
-
MCHR1
-
GPR55
-
Melanocortin Receptor
-
Orexin Receptor
-
Somatostatin Receptor
-
GCGR
-
G protein-coupled Bile Acid Receptor 1
-
GnRH Receptor
-
食欲素受体
-
Prostaglandin Receptor(前列腺素受体)
-
Immunology/Inflammation(免疫学/炎症)
-
COX
-
Interleukin Related
-
SIK(Salt-inducible Kinase)
-
Complement System
-
Aryl Hydrocarbon Receptor(芳烃受体)
-
TLR(Toll-like Receptor)
-
NLR(NOD-like Receptor)
-
PGE synthase
-
SPHK
-
PD-1/PD-L1
-
Thrombopoietin Receptor
-
NOD-like Receptor (NLR)
-
Immunology/Inflammation
-
Cyclic GMP-AMP Synthase
-
FLAP
-
Galectin
-
IFNAR
-
MyD88
-
NO Synthase
-
STING
-
CD73
-
COX
- JAK/STAT Signaling
- MAPK/ERK Pathway
-
Membrane Transporter/Ion Channel(膜转运/离子通道)
-
CFTR
-
Sodium Channel(钠通道)
-
Chloride Channel
-
Calcium Channel(钙通道)
-
NKCC
-
SGLT
-
Na+/K+ ATPase
-
TRP Channel
-
GABA Receptor
-
nAChR
-
iGluR (离子型谷甘酸受体)
-
Na+/Ca2+ Exchanger
-
Potassium Channel(钾通道)
-
URAT1
-
BCRP
-
CRAC Channel
-
Proton Pump
-
P2X Receptor
-
Na+/HCO3- Cotransporter
-
Monoamine Transporter
-
ATP Synthase
-
P-glycoprotein
-
EAAT2
-
GLUT
-
GlyT
-
HCN Channel
-
CRM1
-
CFTR
-
Metabolic Enzyme/Protease(代谢酶/蛋白酶)
-
PDE(Phosphodiesterase)
-
HMGCR(HMG-CoA Reductase)
-
RAR/RXR
-
Ser/Thr Protease
-
Carbonic Anhydrase
-
MMP
-
PGC-1α
-
ROR
-
Dipeptidyl Peptidase
-
Cathepsin
-
Glutathione Peroxidase
-
SCD(Stearoyl-CoA Desaturase)
-
Nampt
-
NEDD8-activating Enzyme
-
Dopamine β-hydroxylase
-
FXR
-
E1/E2/E3 Enzyme
-
Proteasome inhibitor
-
Pyruvate Kinase(丙酮酸激酶)
-
NADPH Oxidase
-
Aminopeptidase(氨基肽酶)
-
Phosphatase
-
Xanthine Oxidase(黄嘌呤氧化酶)
-
IDO
-
HIF/HIF Prolyl-Hydroxylase
-
LXR
-
ROS(Reactive Oxygen Species)
-
Drug Metabolite
-
Endogenous Metabolite
-
ATP Citrate Lyase(ATP柠檬酸裂解酶)
-
FAAH(脂肪酸酰胺水解酶)
-
IDH(异柠檬酸脱氢酶)
-
Acyltransferase
-
Phospholipase(磷脂酶)
-
前列腺素D合成酶(PGDS)
-
乙醛脱氢酶(ALDH)
-
ATGL(脂肪甘油三酯脂酶)
-
Factor Xa(凝血因子Xa)
-
Lipoxygenase
-
Gutathione S-transferase
-
Enolase
-
HIV Protease
-
Acetyl-CoA Carboxylase
-
PAI-1
-
Phosphatase Inhibitor
-
Thrombin
-
COMT
-
Neprilysin
-
Glucokinase
-
Fatty Acid Synthase
-
Dihydroorotate Dehydrogenase
-
CETP
-
Adenosine Deaminase
-
ACE
-
Glutaminase
-
Mitochondrial Metabolism
-
HIV Integrase
-
Aldose Reductase
-
Lactate Dehydrogenase
-
Mineralocorticoid Receptor
-
Aminoacyl-tRNA Synthetase
-
5 alpha Reductase
-
Cytochrome P450
-
Epoxide Hydrolase
-
FABP
-
FASN
-
HMGCR
-
MAGL
-
PDHK
-
Carboxypeptidase
-
Elastase
-
Tryptophan Hydroxylase
-
Phosphoglycerate Dehydrogenase (PHGDH)
-
PDE(Phosphodiesterase)
- Neuronal Signaling(神经信号通路)
- NF-κB信号通路
- PI3K/Akt/mTOR
- PROTAC降解
- Stem Cell/Wnt
- TGF-beta/Smad(信号通路)
- Vitamin D Related
- 其他信号通路
- Antibody-drug Conjugate/ADC Related
- 透析袋
010-60605840